Ecology
The Dutch coast is generally described as mesotidal, barred, dissipative, and moderately exposed (Janssen & Mulder 2005). Based on tidal range and fall velocity of sand, the Holland coast and Southwest coast are considered barred intermediate and the Wadden coast ultra-dissipative. The beach exposure, i.e. dynamics, is of great influence on the occurrence of species (Janssen & Mulder 2004). As tide range or wave energy increases, or sand particle size decreases, beaches become wider, flatter and more dissipative. Faunal communities increase linearly in species richness and exponentially in abundance over this range of beach types (McLachlan 1996). The beaches of the Holland coast and Southwest coast are generally more exposed than the beaches of the Wadden coast.
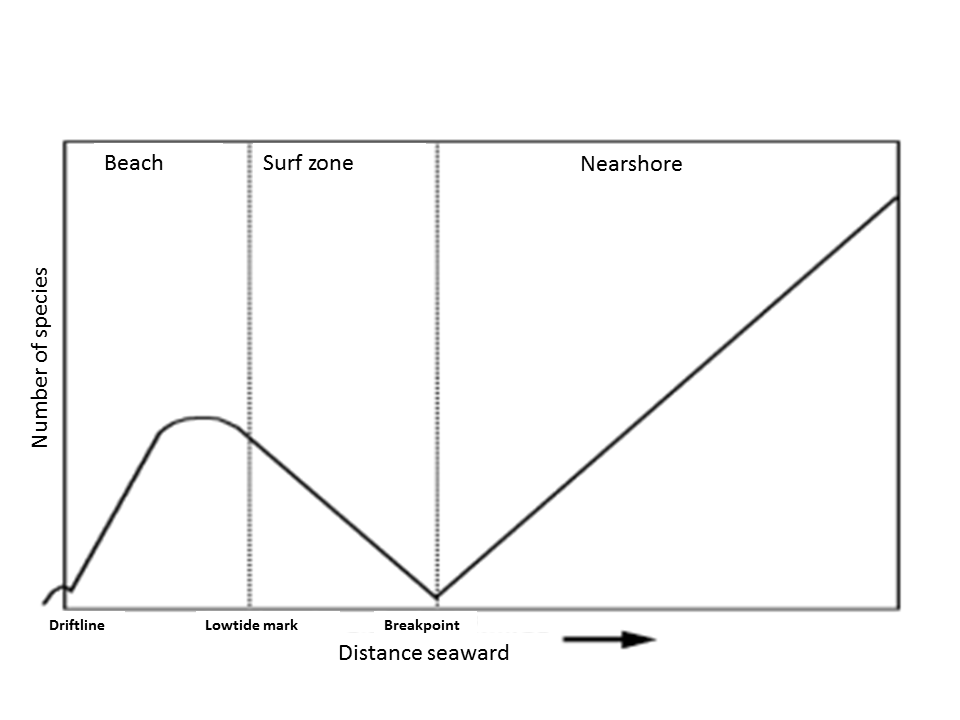
The macrozoobenthos of the beaches and foreshore is a well-investigated group and consitsts of molluscs (bivalves and snails); worms; spiny-skinned animals (Echinodermata); and shrimp-like animals (crustaceans). Different ecological zones can be distinguished on the Dutch sandy coast. Generally, the number of species increases with increasing depth and also correlates with grain size, bed slope and bed morphology.
The beach
The most common species on the beaches of The Netherlands is the bristle worm Scolelepis squamata, an important food source for the Sanderling Calidris alba, which is protected under the EU Birds Directive. Two other characteristic species that are commonly reported are Bathyporeia pilosa – the sand digger shrimp, and Euridice pulchra, the speckled sea louse. The beaches of the Wadden Sea islands are generally characterized by relatively fine sand with low carbonate content and large variation in grain size and a high number of species and densities compared to the mainland coast.
The surf zone and shoreface
The surf zone consists of along shore oriented breaker bars. The number if species is generally low in the surf zone, but total biomass is high (dominated by a few species). In this zone the Polychaetes (worms) are dominant. The crests of the breaker bars have lower species densities probably due to higher wave energy. The troughs between the breaker bars primarily have large numbers of the sand mason. These worms presumably play an important role in holding on to and stabilising the sediment and thus the coast (Janssen & Mulder 2004, Janssen & Mulder 2005). The shoreface, seawards from the outer breaker bank, hosts a lot more species. They represent primarily the main groups Amphipoda, Bivalvia and Echinoïdea. The non-native American razor clam (Ensis directus) is the most abundant species on the Dutch coast (Goudswaard et al. 2008).
Monitoring ecological effects
Possible ecosystem effects of nourishments can be divided into direct and indirect effects. Direct effects are mostly related to burial of benthic species. Indirect effects are caused by change in habitat through the introduction of ‘exotic’ sediment (i.e. sediment from another location with different properties). Altered sediment properties affect the habitat suitability for benthos, such as: level of the seabed; penetrability; organic matter content; grain size; and silt content. For example, impacts at nourished sites in the southeastern United States were observed during monitoring as a result of nearshore turbidity, direct burial of organisms and extreme habitat alterations (Lankford & Baca 1989).
Since the 1980s, several monitoring studies have been conducted on macrobenthos and how it is affected by sand nourishments in the Dutch coastal zone. Some of these involved site-specific projects and some are part of yearly monitoring programs. Baptist et al. (2008) give an overview. Most studies assess short term impacts in and near nourishment sites, quantifying the elimination and early recovery of fauna but few studies cover longer periods. Recovery has only been studied on a limited number of (opportunistic) species. No information is available on the degree of recovery on the level of biological communities.
Survival, migration and recruitment may all contribute to the recovery after disturbance (Van Dalfsen & Essink 2001). The recovery after nourishment therewith depends on many factors, such as the application method/location, the sediment characteristics (influencing both chance of survival and recruitment), the species resistance and resilience and the season of application. Recovery can sometimes be fairly rapid (e.g. some months to <1 year, because of the quick dispersal of sediments and/or the intrinsic tolerance of the assemblages) but can quite often be long-lasting, particularly when the sediments alter the native habitat characteristics, or have high organic loads and/or are highly polluted (Colosio et al. 2007). As the sand used for nourishment of the Dutch coast is not polluted or organically enriched, benthos should be able to recover relatively fast, assuming other sediment characteristics (i.e. grain size and mud content) are fairly similar to the original sediment.
Following shoreface nourishment in Dutch coastal waters a short-term opportunistic response of the benthic community was observed followed by an almost complete recovery of community composition and structure after four years (Van Dalfsen & Essink 2001). Opportunistic species will quickly recolonise the affected site, but long-living bivalve species or some sea urchins (such as the sea potato, Echinocardium cordatum) do not reproduce each year. In general, soft bottom benthic communities show partial recovery in one year and full recovery from 18 to 24 months (Allen & Hardy 1980) up to a maximum of 5 years (Mulder et al. 2005).